
ГЛУБОКОВОДНЫЕ ЗОНЫ
Глубоководные (абиссальные) зоны - области океана глубиной больше 2000 м - занимают больше половины поверхности земли. Следственно, это особенно распространенная среда обитания, но она же остается и наименее изученной. Только в последнее время, вследствие происхождению глубоководных агрегатов, мы начинаем познавать данный чудесный мир.
Для глубинных зон характерны непрерывные данные: холод, темнота, большое давление (больше 1000 атмосфер), из-за непрерывной циркуляции воды в глубоководных морских течениях там нет недостатка кислорода. Эти зоны существуют в течение дюже длинного времени, там нет барьеров для распространения организмов.
В полной темноте нелегко обнаружить пищу либо компаньона, следственно обитатели морских глубин приспособились узнавать друг друга с поддержкой химических сигналов; некоторые глубоководные рыбы владеют биолюминесцентными органами, в которых содержатся светящиеся бактерии-симбионты. Глубоководные рыбы - удильщики пошли дальше: когда самец (больше мелкий) находит самку, он прикрепляется к ней и у них становится всеобщим даже кровообращение. Другое последствие темноты - неимение фотосинтетических организмов, следственно, сообщества получают питательные вещества и энергию из усопших организмов, попадающих на морское дно. Это могут быть как громадные киты, так и микроскопичный планктон. Мелкие частицы зачастую образуют хлопья «морского снега», смешиваясь со слизью, питательными веществами, бактериями и простейшими. По пути на дно огромная часть органического материала съедается либо из него выдается много азота, следственно к тому времени, когда остатки заканчивают свой путь, они становятся не дюже питательными. Это одна из причин, по которым насыщенность биомассы на морском дне дюже мала.
Главным объектом грядущих изысканий глубоководных зон должна стать роль бактерий в пищевой цепи.
См. также статью «Океаны».
Из книги Сон - тайны и парадоксыавтораВейн Александр МоисеевичГипногенные зоны В предыдущей главе мы нарисовали внешнюю картину сна. Если не считать такие явления, как сомнамбулизм и броски-раскачивания, картина эта отлично знакома всякому. Сейчас перед нами стоит больше трудная задача - представить себе, что происходит во время сна
Из книги Всеобщая экологияавтора Чернова Нина Михайловна 4.1.1. Экологические зоны Мирового океана В океане и входящих в него морях различают раньше каждого две экологические области: толщу воды - пелагиаль и дно - бенталь (рис. 38). В зависимости от глубины бенталь делится на сублиторальную зону - область плавного понижения суши
Из книги Жизнеобеспечение экипажей летательных агрегатов позже вынужденного приземления либо приводнения (без иллюстраций)автораВолович Виталий Георгиевич
Из книги Жизнеобеспечение экипажей летательных агрегатов позже вынужденного приземления либо приводнения [с иллюстрациями]автораВолович Виталий Георгиевич
- Вводный урокбесплатно;
- Большое число опытных преподавателей (нейтивов и русскоязычных);
- Курсы НЕ на определенный срок (месяц, полгода, год), а на определенное число занятий (5, 10, 20, 50);
- Более 10 000 довольных заказчиков.
- Стоимость одного занятия с русскоязычным преподавателем -от 600 рублей, с носителем языка -от 1500 рублей
Экологические области Мирового океана, - области (зоны) океанов, где систематический состав и разделение морфологических и физиологических особенностей морских организмов узко связаны с окружающими их условиями среды: пищевыми источниками, температурным, солевым, световым и газовым режимом водных масс, другими их физическими и химическими свойствами, физическими и химическими свойствами морских грунтов и, наконец, с другими организмами, населяющими океаны и образующими совместно с ними биогеоценотические системы. Все перечисленные свойства испытывают существенные метаморфозы от поверхностных слоев в глубины, от побережий к центральным частям океана. В соответствии с указанными абиотическими и биотическими факторами среды в океане выдаются экологические зоны, а организмы делятся на экологические группы.
Все живые организмы океана в совокупности делятся набентос, планктон и нектон.Первая группа включает организмы, живущие на дне в прикреплённом либо свободноподвижном состоянии. Это в большинстве огромные организмы, с одной стороны, многоклеточные водоросли (фитобентос), а с иной - разные звериные: моллюски, черви, ракообразные, иглокожие, губки, кишечнополостные и др. (зообентос).Планктонсостоит в большинстве из мелких растительных (фитопланктон) и животных (зоопланктон) организмов, находящихся во взвешенном состоянии в воде и носящихся совместно с нею, органы движения у них слабые.Нектон- это общность животных организмов, обыкновенно больших размеров, владеющих крепкими органами передвижения, - морские млекопитающие, рыбы, головоногие моллюски-кальмары. Помимо этих трёх экологических групп, дозволено выделить плейстон и гипонейстон.
Плейстон- общность организмов, существующих в самой поверхностной плёнке воды, часть их тела погружена в воду, а часть выставляется над поверхностью воды и исполняет роль паруса.Гипонейстон- организмы поверхности водного слоя в несколько сантиметров, Всей жизненной форме свойственны определённая форма тела и некоторые придаточные образования. Нектонным организмам свойственна торпедообразная форма тела, планктонным - приспособления к парению (шипы и отростки, а также газовые пузыри либо капли жира, сокращающие вес тела), защитные образования в виде панцирей, скелетов, раковин и т. п.
Главнейшим фактором в разделении морских организмов является разделение пищевых источников как поступающих с побережий, так и создаваемых в самом водоёме. По методу питания морские организмы могут быть распределены на хищников, растительноядных, фильтраторов - сестонофагов (сестон - это взвешенные в воде мелкие организмы, органический детрит и минеральная взвесь), детритофагов и грунтоедов.
Как и во каждом ином водоёме, живые организмы океана могут быть поделены на продуцентов, консументов (покупателей) и редуцентов (возвращающих обратно). Основная масса нового органического вещества создаётся продуцентами-фотосинтетиками, способными существовать только в верхней зоне, довольно отменно освещаемой ясными лучами и не простирающейся глубже 200 м, впрочем основная масса растений приурочена к верхнему слою воды в несколько десятков метров. У побережий - это многоклеточные водоросли: макрофиты (зелёные, бурые и красные), растущие в прикреплённом ко дну состоянии (фукусы, ламинарии, алярии, саргассы, филлофора, ульва и мн. др.), и некоторые цветковые растения (зостера филлоспадикс и др.). Иная масса продуцентов (одноклеточные планктонные водоросли, основным образом диатомовые и перидиниевые) во множестве населяет поверхностные слои моря. Консументы существуют за счёт готовых органических веществ, сделанных продуцентами. Это каждая масса животных, населяющих моря и океаны. Редуценты - это мир микробов, разлагающих органические соединения до самых примитивных форм и опять создающих из этих последних больше трудные соединения, нужные растительным организмам для их жизнедеятельности. В какой-то степени микробы являются также хемосинтетиками - они продуцируют органическое вещество, переводя одни химические соединения в другие. Так совершаются циклические процессы органических веществ и жизни в морских водоёмах.
По физическим и химическим особенностям водной массы океана и по рельефу дна его подразделяют на несколько вертикальных зон, которым свойственны определённый состав и экологические особенности растительного и звериного населения (см. схему). В океане и входящих в него морях различают раньше каждого две экологические области: толщу воды -пелагиальи дно -бенталь.В зависимости от глубиныбентальделится насублиторальнуюзону - область плавного понижения суши до глубины приблизительно 200 м,батиальную- область крутого склона иабиссальную зону- область океанического ложа со средней глубиной 3-6 км. Еще больше большие области бентали, соответствующие впадинам океанического ложа, называютультраабиссалью.Кромка берега, заливаемая во время приливов, называетсялиторалью.Выше яруса приливов часть берега, увлажняемая брызгами прибоя, получила названиесупралиторали.
Бентос обитает в самом верхнем горизонте - в литорали. Морская флора и фауна богато заселяют литоральную зону и вырабатывают в связи с этим ряд экологических приспособлений к переживанию периодической осушки, Некоторые звериные плотно закрывают свои домики и раковины, другие зарываются в грунт, третьи забиваются под камни и водоросли либо плотно сжимаются в комок и выделяют на поверхности слизь, препятствующую высыханию. Некоторые организмы выбираются ещё выше самой верхней границы прилива и довольствуются заплесками волн, орошающими их морской водой. Это супралиторальная зона. В состав литоральной фауны входят примерно все огромные группы животных: губки, гидроиды, черви, мшанки, моллюски, ракообразные, иглокожие и даже рыбы, в супралитораль выбираются некоторые водоросли и ракообразные. Ниже самой низкой границы отлива (до глубины около 200 м) простирается сублитораль, либо материковый шельф. По обилию жизни литораль и сублитораль стоят на первом месте, исключительно в умеренной зоне - большие заросли макрофитов (фукусы и ламинарии), собрания моллюсков, червей, ракообразных и иглокожих служат обильным кормом рыбам. Плотность жизни на литорали и сублиторали достигает нескольких килограммов, а изредка десятков килограммов, основным образом за счёт водорослей, моллюсков и червей. Сублитораль - основная область применения человеком сырьевых источников моря - водорослей, беспозвоночных и рыб. Ниже сублиторали располагается батиаль, либо материковый склон, переходящий на глубине 2500-3000 м (по иным данным 2000 м) в ложе океана, либо абиссаль, в свою очередь, подразделяющуюся на подзоны верхнеабиссальную (до 3500 м) и нижнеабиссальную (до 6000 м). В пределах батиали плотность жизни круто падает до десятков грамм и нескольких грамм на 1 м3, а в абиссали до нескольких сотен и даже десятков мг на 1 л3. Наибольшая часть ложа океана занята глубинами 4000-6000 м. Глубоководные впадины с их наибольшими глубинами до 11000 м занимают каждого около 1% площади дна, это ультраабиссальная зона. От побережий до наибольших глубин океана уменьшается не только плотность жизни, но и её многообразие: в поверхностной зоне океана обитают многие десятки тысяч видов растений и животных, а для ультраабиссали знаменито каждого несколько десятков видов животных.
Пелагиальтакже делят на вертикальные зоны, соответствующие по глубине зонам бентали:эпипелагиаль, батипелагиаль, абиссопелагиаль.Нижняя граница эпипелагиали (не больше 200 м) определяется проникновением ясного света в числе, довольном для фотосинтеза. Организмы, обитающие в толще воды, либо пелагиали, относятся кпелагосу.Подобно донной фауне, плотность планктона также испытывает количественные метаморфозы от побережий в центр, части океанов и от поверхности в глубины. У побережий плотность планктона определяется сотнями мг на 1 л3, изредка несколькими граммами, а в средних частях океанов несколькими десятками граммами. В глубинах океана она падает до нескольких мг либо долей мг в 1 м3. Растительный и звериный мир океана с увеличением глубины претерпевает закономерные метаморфозы. Растения обитают только в верхней 200-метровой толще воды. Прибрежные макрофиты в своём приспособлении к нраву освещения испытывают смену состава: самые верхние горизонты заняты предпочтительно зелёными водорослями, после этого идут бурые и глубже каждого проникают красные водоросли. Это связано с тем, что в воде стремительней каждого затухают красные лучи спектра, а глубже каждого идут синие и фиолетовые лучи. Растения окрашиваются в добавочный цвет, что обеспечивает наилучшие данные фотосинтеза. Такая же смена окраски отслеживается и у донных животных: на литорали и сублиторали они предпочтительно серых и бурых оттенков, а с глубиной всё огромнее проявляется красная окраска, но рациональность этой смены цвета в данном случае иная: окраска в добавочный цвет делает их заметными и охраняет от недругов. У пелагических организмов и в эпипелагиали и глубже отслеживается потеря пигментации, некоторые звериные, исключительно кишечнополостные, становятся прозрачными, как стекло. В самом поверхностном слое моря прозрачность содействует прохождению через их тело ясных лучей без пагубного воздействия на их органы и ткани (исключительно в тропиках). Помимо того, прозрачность тела делает их заметными и выручает от недругов. Наравне с этим с глубиной некоторые планктонные организмы, исключительно ракообразные, приобретают красную окраску, что при слабом освещении делает их заметными. Не подчиняются этому правилу глубоководные рыбы, в большинстве они окрашены в чёрный цвет, правда и среди них имеются депигментированные формы.
Земная кора бывает материковая и океаническая. Материковая - это суша и на ней есть горы, равнины и низменности - их видно и неизменно дозволено по ним пройти. А вот какой бывает океаническая земная кора мы узнаем из темы “Дно мирового океана” (6 класс).
Изучение океанического дна
Первыми, кто стал постигать мировой океан, стали британцы. На военном корабле “Chellenger” под командованием Джоржа Нэйса, они прошли всю акваторию мира и собрали много пригодной информации, которую ученые классифицировали еще 20 лет. Они измеряли температуру воды, животных, но самое главное - они первые определили строение дна океанов.
Прибор, которым постигают глубину, именуется эхолот. Он размещен в нижней части корабля и периодично посылает сигнал такой силы, дабы он мог достичь дна, отразится и возвратиться на поверхность. Согласно законам физики, звук в воде движется со скоростью 1500 м. за секунду. Таким образом, если звук возвратился за 4 секунды, то дна он добился теснее на 2-й, и глубина в этом месте равна 3000 м.
Как выглядит земля под водой?
Ученые выделяют основные части дна мирового океана:
- Подводная окраина континентов;
- Переходная зона;
- Ложе океана.
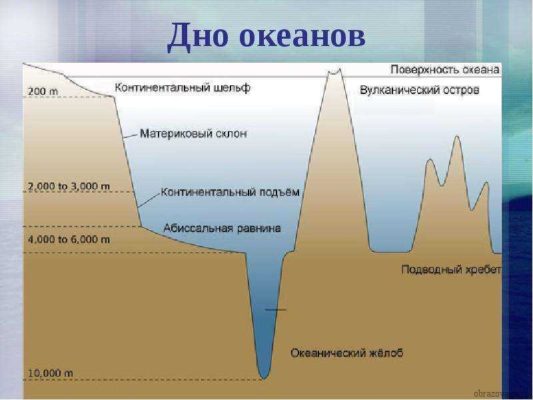
Рис. 1. Рельеф дна мирового океана
Континент неизменно отчасти уходит под воду, следственно подводная окраина разделяется на материковый шельф и материковый склон. Фраза “выйти в открытое море” обозначает оставить рубеж материкового шельфа и склона.
Материковая отмель (шельф) - это часть суши, погруженная под воду на глубину до 200 м. На карте она выделена бледно-голубым либо белым цветом. Крупнейший шельф - в северных морях и на Северно-Ледовитом океане. Минимальный - в Северной и Южной Америке.
ТОП-2 статьикоторые читают совместно с этой
Материковая отмель отлично прогревается, следственно это основная зона для курортов, хозяйств по добыче и разведению морепродуктов. В этой части океана добывают нефть
Материковый склон формирует границы океанов. Материковый склон считается от края шельфа и до глубины в 2 километра. Если бы склон был на суше, то это бы был высоченный обрыв с дюже резкими, примерно прямыми склонами. Но помимо своей крутизны, в них таится еще одна угроза - океанические желоба. Это тесные ущелья, уходящие под воду еще на тысячи метров. Самым огромным и знаменитым желобом считается Марианская впадина.
Ложе океана
Там, где кончается материковый уступ, начинается ложе океана. Это основная его часть, где существуют глубоководные котловины (4 - 7 тыс. м.) и возвышенности. Ложе океана размещается на глубине от 2 до 6 км. Звериный мир представлен дюже слабо, от того что в этой части фактически нет света и дюже холодно.

Рис. 2. Изображение дна океана
Значимое место занимают срединно океанические хребты. Они представляют собой крупную горную систему, как на суше, только под водой, простирающиеся по каждого океана. Всеобщая протяженность хребтов - около 70 000 км. Они имеют свою трудную конструкцию: ущелья и большие склоны.
Хребты образуются на стыках литосферных плит и являются источниками вулканов и землетрясений. Некоторые острова имеют дюже увлекательное происхождение. В тех местах, где скапливалась вулканическая порода и в результате вышла на поверхность, образовался остров Исландия. Именно следственно тут много гейзеров и жгучих источников, а сама страна представляет собой неповторимый природный заповедник.
Рис. 3. Рельеф Атлантического океана
Океаническое дно
Почва океана представляет собой морские осадки. Они бывают 2-х типов: материковые и океанические. Первые произошли с суши: галька, песок, другие частицы с берега. Вторые - это донные отложения, сформировавшиеся океаном. Это остатки морских обитателей, вулканический пепел.
Что мы узнали?
Строение дна океана дюже неравномерное. Выделяют три основные его части: материковая окраина (делящаяся на материковый шельф и склон), переходная зона и ложе океана. Именно в центральной его части образовался ошеломительный рельеф - срединно-океанический хребет, представляющий цельную горную систему, опоясывающую фактически всю Землю.
Тест по теме
Оценка доклада
Средняя оценка:4.2 . Каждого получено оценок: 100.
Зона эвфотическая - верхняя (в среднем 200 м) зона океана, где освещенность довольна для фото-синтетической жизнедеятельности растений. Тут богато представлен фитопланктон. Особенно насыщенно процесс фотосинтеза идет на глубинах 25-30 м, где освещенность составляет не менее 1/3 от освещенности поверхности моря. На глубине больше 100 м интенсивность освещения снижается до величины 1/100. В районах Мирового океана, где воды исключительно прозрачны, фитопланктон может обитать на глубинах до 150-200 м.
Глубинные воды Мирового океана отличаются крупной однородностью, но совместно с тем все типы этих вод имеют свои характерные черты. Формируются глубинные воды основным образом в высоких широтах в итоге смешения поверхностных и промежуточных вод в областях циклонических круговоротов, расположенных возле континентов. К основным очагам образования глубинных вод относятся северо-западные районы Тихого, Атлантического океанов и районы Антарктиды. Они располагаются между промежуточными и придонными водами. Толщина этих вод в среднем 2000-2500 м. Она максимальна (до 3000 м) в экваториальной зоне и в районе субантарктических котловин.
Глубина D получила наименование глубины трения. На горизонте, равном удвоенной глубине трения, направления векторов скорости дрейфового течения на этой глубине и на поверхности океана совпадут. Если глубина водоема в рассматриваемом районе огромнее глубины трения, то такой водоем следует считать безгранично глубоким. Таким образом, в приэкваториальной зоне Мирового океана глубины, вне зависимости от их реального значения, следует считать малыми и рассматривать дрейфовые течения как течения в неглубоком море.
С глубиной плотность изменяется в связи с изменением температуры, солености и давления. При понижении температуры и увеличении солености плотность возрастает. Впрочем типичная стратификация плотности нарушается в отдельных районах Мирового океана в связи с территориальными, сезонными и другими изменениями температуры и солености. В экваториальной зоне, где поверхностные воды касательно опреснены и имеют температуру 25-28° С, они подстилаются больше солеными холодными водами, следственно плотность круто повышается до горизонта 200 м, а после этого медлительно возрастает к 1500 м, позже чего становится примерно непрерывной. В умеренных широтах, где в предзимнее время происходит охлаждение поверхностных вод, плотность возрастает, прогрессируют конвективные токи и больше плотная вода опускается, а менее плотная подымается к поверхности - появляется вертикальное перемешивание слоев.
В рифтовых зонах Мирового океана было выявлено около 139 глубинных гидротермальных полей (65 из них энергичных, см. рис. 5.1). Дозволено ждать, что число таких систем будет возрастать по мере дальнейших изысканий рифтовых зон. Присутствие 17 энергичных гидротермальных систем по отрезка неовулканической зоны длиной 250 км в рифтовой системе Исландии и по крайней мере 14 энергичных гидротермальных систем по отрезка длиной 900 км в Красном море указывает пространственный диапазон в разделении гидротермальных полей между 15 и 64 км.
Оригинальной зоной Мирового океана, характеризующейся высокой рыбопродуктивностью, является апвеллинг, т.е. подъем вод из глубины в верхние слои океана, как водится, на западных берегах контингентов.
Поверхностная зона (с нижней рубежом на глубине в среднем 200 м) характеризуется высокой динамичностью и изменяемостью свойств вод, обусловленной сезонными колебаниями температуры и ветровым волнением. Объем заключенной в ней воды 68,4 млн. км3, что составляет 5,1% объема воды Мирового океана.
Промежуточная зона (200-2000 м) отличается сменой поверхностной циркуляции с ее широтным переносом вещества и энергии на глубинную, в которой превалирует меридиональный перенос. В высоких широтах к этой зоне приурочен слой больше теплой воды, проникшей из низких широт. Объем воды в промежуточной зоне 414,2 млн. км3, либо 31,0% Мирового океана.
Самая верхняя часть океана, куда проникает свет и где создается первичная продукция, именуется эвфотической. Ее мощность в открытом океане доходит до 200 м, а в прибрежной части - не больше 30 м. По сопоставлению с километровыми глубинами это зона довольно тонкая и отделяется компенсационной зоной от гораздо большей водной толщи, вплотную до самого дна - афотической зоны.
В пределах открытого океана выделяют три зоны, основным отличием которых является глубина проникновения ясных лучей (рис. 6.11).
Помимо экваториальной зоны апвеллинга, подъем глубинных вод появляется там, где крепкий непрерывный ветер отгоняет поверхностные слои от берега огромных водоемов. Рассматривая итоги теории Экмана, дозволено констатировать, что апвеллииг происходит при касательном к храню направлении ветра (рис. 7.17). Смена направления ветра на противоположное ведет к смене апвеллинга на даунвеллинг либо напротив. На зоны апвеллинга доводится каждого 0,1% площади Мирового океана.
Глубоководные рифтовые зоны океана находятся на глубине около 3000 м и больше. Данные жизни в экосистемах глубоководных рифтовых зон дюже оригинальны. Это полная темнота, громадное давление, пониженная температура воды, недочет пищевых источников, высокая насыщенность сероводорода и ядовитых металлов, встречаются выходы жгучих подземных вод, и т. д. В итоге живущие тут организмы претерпели следующие адаптации: редукция плавательного пузыря у рыб либо заполнение его полости жировой тканью, атрофирование органов зрения, становление органов светосвечения и др. Живые организмы представлены гигантскими червями (погонофорами), огромными двустворчатыми моллюсками, креветками, крабами и отдельными видами рыб. Продуцентами выступают сероводородные бактерии, живущие в симбиозе с моллюсками.
Материковый склон - это зона перехода от континентов к ложу океана, расположенная в пределах 200-2440 м (2500 м). Она характеризуется резкой сменой глубин и существенными уклонами дна. Средние уклоны дна 4-7°, в отдельных районах доходят до 13-14°, как, скажем, в Бискайском заливе; знамениты еще огромные уклоны дна вблизи коралловых и вулканических островов.
При подъеме по разломной зоне с раздвижением до глубин 10 км и менее (от яруса дна океана), что примерно соответствует расположению границы Мохоровичича в океанической литосфере, ультра-основная мантийная интрузия может попадать в зону циркуляции термальных вод. Тут при Т= 3 00- 500°С создаются благоприятные данные для процесса серпентинизации ультрабазитов. Наши расчеты (см. рис. 3.17, а), а также отслеживаемые над такими разломиыми зонами повышенные значения теплового потока (в 2-4 раза превышающие типичные значения q для океанической коры) полагают присутствие температурного промежутка серпентинизации на глубинах 3-10 км (эти глубины мощно зависят от расположения кровли высокотемпературного интрузивного мантийного материала). Постепенная серпентинизация перидотитов понижает их плотность до значений, меньших плотности окружающих пород океанической коры, и приводит к увеличению их объема на 15-20%.
В будущем будет видно, что глубина трения в средних широтах и при средних скоростях ветра бывает невелика (приблизительно порядка 100 м). Следственно, уравнения (52) дозволено использовать в примитивный форме (47) во каждом море со сколько-нибудь существенной глубиной. Исключение составляет область мирового океана, лежащая по соседству с экватором, где ?sin ф тяготится к нулю, а глубина трения - к бесконечности. Разумеется, пока тут речь идет об открытом море; что касается прибрежной зоны, то о ней придется много говорить в будущем.
Батиаль (от греч. - большой)- зона, занимающая промежуточное расположение между материковой отмелью и ложем океана (от 200-500 до 3000 м), т. е. соответствует глубинам материкового склона. Эта экологическая область характеризуется стремительным нарастанием глубины и гидростатистическим давлением, постепенным понижением температуры (в низких и средних широтах- 5-15°С, в высоких- от 3° до- 1°С), отсутствием фотосинтезирующих растений и др. Донные осадки представлены органогенными илами (из скелетных остатков фораминифер, кокколитофорид и пр.). В этих водах буйно прогрессируют автотрофные хемосинтезирующие бактерии; характерны многие виды плеченогих, морские перья, иглокожие, десятиногие ракообразные, из придонных рыб обыкновенны долгохвосты, угольная рыба и др. Биомасса - традиционно граммы, изредка десятки граммов/м2.
От описанных выше сейсмоактивных зон срединно-океанических хребтов значительно отличаются те, которые расположены в районах островных дуг и энергичных континентальных окраин обрамления Тихого океана. Отменно знаменито, что характерная специфика таких зон - их проникновение до дюже огромных глубин. Глубины очагов землетрясений тут достигают 600 и больше километров. При этом, как показали изыскания С. А. Федотова, Л. Р. Сайкса и А. Хасегавы, ширина уходящей в глубь зоны сейсмической активности не превышает 50-60 км. Иная значимая отличительная специфика этих сейсмоактивных зон - механизмы в очагах землетрясений, абсолютно определенно свидетельствующие о сжатии литосферы в районе внешнего края островных дуг и энергичных континентальных окраин.
Экосистема глубоководных рифтовых зон океана - эта неповторимая экосистема была открыта заокеанскими учеными в 1977 г. в рифтовой зоне подводного хребта Тихого океана. Тут на глубине 2 600 м, в сплошной темноте, при обильном оглавлении сероводорода и ядовитых металлов, выдающихся из гидротермальных источников, были найдены «оазисы жизни». Живые организмы были представлены гигантскими (до 1-1,5 м длиной), живущими в трубках червями (погонофорами), огромными белыми двустворчатыми моллюсками, креветками, крабами и отдельными экземплярами оригинальных рыб. Биомасса только погонофор достигала 10-15 кг/м2 (на соседних участках дна - каждого 0,1-10 г/м2). На рис. 97 показаны особенности данной экосистемы в сопоставлении с наземными биоценозами. Серобактерии составляют первое звено пищевой цепи этой уникальной экосистемы, дальше идут погонофоры, внутри тела которых обитают бактерии, перерабатывающие сероводород в нужные питательные вещества. В экосистеме рифтовых зон 75 % биомассы составляют организмы, живущие в симбиозе с хемоавтотрофными бактериями. Хищники представлены крабами, брюхоногими моллюсками, отдельными видами рыб (макруридами). Схожие «оазисы жизни» были найдены в глубоководных рифтовых зонах многих районов Мирового океана. Подробнее дозволено ознакомиться в книге французского ученого Л. Лобье «Оазисы на дне океана» (Л., 1990).
На рис. 30 показаны основные экологические зоны Мирового океана, показывающие вертикальную зональность разделения живых организмов. В океане раньше каждого выделяют две экологические области: толщу воды - пелагиалъ и дно - ёенталъ. В зависимости от глубины бенталь делится на литоральную (до 200 м), батиальную (до 2500 м), абиссальную (до 6000 м) и улътраабис-сальную (глубже 6000 м) зоны. Пелагиаль также подразделяется на вертикальнее зоны, соответствующие по глубине зонам бентали: эпипелаги-аль, батипелагиаль и абиссопелагиалъ.
Резкий материковый склон океана заселен представителями батиальной (до 6000 м), абиссальной и ультраабиссальной фауны; в этих зонах, за пределами доступного для фотосинтеза освещения, растения отсутствуют.
Абиссаль (от греч. - бездонный) - экологическая зона разделения жизни на дне Мирового океана, соответствующая глубинам океанического ложа (2500-6000 м).
До сего времени речь шла о воздействии на физические параметр: океана и лишь неявно предполагалось, что таким образом чере эти параметры идет влияние и на экосистемы. С одной стс роны, подъем богатых биогенными солями глубинных вод може служить фактором возрастания биопродуктивности этих в оС щем-то бедных районов. Можн рассчитывать на то, что подъем глубинных вод дозволит снизит температуру поверхностных вод правда бы в каких-то локальны зонах с одновременным увеличением за счет возрастания раствс римости кислорода оглавления последнего. С иной стороны, со сбросом в среду холодны вод связана смерть теплолюбивых видов с низкой термическо стабильностью, метаморфоза видового состава организмов, кормово базы и т. п. Помимо того, экосистема будет непрерывно подвергатьс воздействию биоцидов, препятствующих обрастанию рабочих эле ментов станции, воздействию разных реагентов, металлов, шг сел и других побочных выбросов.
Основным фактором, дифференцирующим морскую биоту, является глубина моря (см. рис. 7.4): материковый шельф круто сменяется материковым склоном, плавно переходящим в материковое подножие, которое опускается ниже к ровному ложу океана - абиссальной равнине. Этим морфологическим частям океана приблизительно соответствуют следующие зоны: неритическая - шельфу (с литоралью - приливно-отливной зоной), батиальная - материковому склону и его подножию; абиссальная - область океанических глубин от 2000 до 5000 м. Абиссальная область разрезается большими впадинами и ущельями, глубина которых больше 6000 м. Область открытого океана за пределами шельфа называют океанической. Все население океана, так же как и в пресноводных экосистемах, делится п планктон, нектон, бентос. Планктон и нектон, т.е. все, что живет в открытых водах, образует так называемую пелагическую зону.
Принято считать, что береговые станции рентабельны, если требуемые глубины с подходящей температурой охлаждающей воды находятся довольно близко от побережья и длина трубопровода не превышает 1-3 км. Такая обстановка характерна для многих островов тропического пояса, представляющих собой вершины подводных гор и потухших вулканов и не имеющих присущного континентам протяженного шельфа: их берега довольно резко спускаются по направлению к океанскому ложу. Если берег довольно удален от зон требуемых глубин (скажем, на островах, окруженных коралловыми рифами) либо отделен полого понижающимся шельфом, то для сокращения длины трубопроводов энергоблоки станций могут быть вынесены на неестественные острова либо стационарные платформы - аналоги используемых при морской добыче нефти и газа. Превосходства наземных и даже островных станций в том, что пропадает надобность в создании и обслуживании дорогостоящих, подвергающихся воздействию открытого океана сооружений - будь то неестественные острова либо стационарные основания. Впрочем два значительных фактора, ограничивающих береговое базирование, все-таки остаются: органиченность соответствующих островных территорий и надобность прокладки и охраны трубопроводов.
Впервой морфологическая колляция и типизация океанических разломных зон по морфологическим знакам (на примере разломов северо-восточной части Тихого океана) была сделана Г.Менардом и Т.Чейсом. Они определили разломы как ’’длинные и тесные зоны мощно расчлененного рельефа, характеризующиеся присутствием вулканов, линейных хребтов, уступов и обыкновенно отделяющие друг от друга разные топографические глубинке с неодинаковыми территориальными глубинами”. Выраженность трансформных разломов в рельефе дна океана и аномальных геофизических полях, как водится, достаточно резкая и отчетливая. Это удостоверили и бесчисленные детальные изыскания, проведенные в последние годы. Высокие приразломные хребты и большие впадины, сбросы и трещины характерны для зон трансформных разломов. Аномалии А, АТ, теплового потока и другие свидетельствуют о гетерогенности строения литосферы и трудной динамике разломных зон. Помимо того, разновозрастные блоки литосферы, расположенные по различные стороны от разлома, в соответствии с законом V/ имеют разное строение, выражающееся в разных глубине дна и толщине литосферы, что создает добавочные территориальные аномалии в геофизических полях.
Область континентального шельфа, неритическая область, если ее площадь ограничить глубиной до 200 м, составляет около восьми процентов площади океана (29 млн км2) и является самой богатой в фау-нистическом отношении в океане. Прибрежная зона благоприятна по условиям питания, даже в дождевых тропических лесах нет такого многообразия жизни, как тут. Дюже богат кормом планктон за счет личинок бентосной фауны. Личинки, которые остаются несъеденны-ми, оседают на субстрат и образуют либо эпифауну (прикрепленную), либо инфауну (закапывающуюся).
У планктона также выражена вертикальная дифференциация при адаптации различных видов к различным глубинам и различной интенсивности освещения. Вертикальные миграции влияют на разделение этих видов, и потому ярусность по вертикали в этом сообществе менее очевидна, чем в лесу. Сообщества освещенных зон на дне океана ниже яруса прилива дифференцированы отчасти интенсивностью света. Виды зеленых водорослей концентрируются на мелководье, виды бурых водорослей распространены на несколько огромных глубинах, а еще ниже исключительно обильны красные водоросли. Бурые и красные водоросли содержат, помимо хлорофилла и каротиноидов, добавочные пигменты, что разрешает им применять свет низкой интенсивности и чудесный по спектральному составу от света в мелководьях. Вертикальная дифференциация, таким образом, является всеобщей чертой природных сообществ.
Абиссальные ландшафты - царство тумана, холодных, малоподвижных вод и дюже бедной органической жизни. В олиштроф-ных зонах Океана биомасса бентоса колеблется от 0,05 и менее до 0,1 г/м2, несколько возрастая в областях богатого поверхностного планктона. Но и тут, на таких крупных глубинах, встречены «оазисы жизни». Почвы» абиссальных ландшафтов образованы илами. Состав их, как и наземных почв, зависит от широты места и высоты (в данном случае глубины). Где-то на глубине 4000-5000 м преобладавшие ранее карбонатные илы сменяются бескарбонатными (красными глинами, радио-ляриевым илом в тропиках и диатомовым в умеренных широтах).
Тут х показатель термической диффузии пород литосферы, Ф - функция вероятности, (Т + Сг) - температуры мантии под осевой зоной срединного хребта, т.е. при / = 0. В модели пограничного слоя глубина изотерм и подошвы литосферы, а также глубина дна океана И, отсчитываемая от ее значения на оси хребта, возрастают пропорционально значению V/.
В высоких широтах (выше 50°) происходит уничтожение сезонного термоклина с конвективным перемешиванием водных масс. В приполярных областях океана имеет место восходящее движение глубинных масс. Следственно эти широты океана относятся к высокопродуктивным районам. По мере последующего движения к полюсам производительность начинает падать из-за понижения температуры воды и уменьшения ее освещенности. Для океана характерна не только пространственная изменяемость производительности, но и повсеместная сезонная изменяемость. Сезонная изменяемость производительности обусловлена в существенной мере реакцией фитопланктона на сезонные метаморфозы условий среды обитания, раньше каждого освещенности и температуры. Наибольшая сезонная контрастность отслеживается в умеренной зоне океана.
Поступление магмы в магматическую камеру происходит, видимо, эпизодически, и является функцией высвобождения большого числа расплавленного вещества с глубин больше 30 - 40 км в верхней мантии. Насыщенность расплавленного вещества в центральной части сегмента приводит к увеличению объема (разбуханию) магматической камеры и миграции расплава по оси к краям сегмента. С приближением к трансформному разлому глубина кровли, как водится, опускается вплотную до полного исчезновения соответствующего горизонта возле трансформного разлома. Это в существенной степени обусловлено охлаждающим влиянием больше ветхого литосферного блока, граничащего с осевой зоной по трансформному разлому (результат трансформного разлома). Соответственно, отслеживается и постепенное погружение яруса дна океана (см. рис.3.2).
В антарктическом районе южного полушария дно океанов покрыто ледниковыми и айсберговыми отложениями и диатомовыми илами, встречающимися также на севере Тихого океана. Дно Индийского океана выстлано илом с огромным оглавлением углекислого кальция; глубоководные впадины - красной глиной. Особенно разновидны отложения дна Тихого океана, где на севере преобладают диатомовые илы, северная половина покрыта в области глубин свыше 4000 м красной глиной; в приэкваториальной зоне восточной части океана распространены илы с кремнистым остатком (радиоляриевые), в южной половине на глубинах до 4000 м встречаются известково-карбонатные илы,. красная глина, на юге - диатомовые и ледниковые отложения. В районах вулканических островов и коралловых рифов встречается вулканический и коралловый песок и ил (рис. 7).
Смена континентальной земной коры на океаническую происходит не потихоньку, а скачкообразно, сопровождаясь образованием морфоструктур специального рода, присущных переходным, вернее- контактным, зонам. Изредка их называют периферическими областями океанов. Главнейшими морфоструктурами их являются островные дуги с действующими вулканами, круто переходящие в сторону океана в глубоководные желоба. Именно тут, в тесных, глубочайших (до 11 км) впадинах Мирового океана, проходит структурная граница континентальной и океанической коры, совпадающая с глубинными разломами, знаменитыми у геологов под наименованием зоны Заварицкого - Бенъофа. Разломы, падающие под континент, идут на глубину до 700 км.
2-й особый эксперимент по постижению синоптической изменяемости океанских течений («Полигон-70») был проведен советскими океанологами во главе с Университетом океанологии АН СССР в феврале-сентябре 1970 г. в северной пассатной зоне Атлантики, где в течение шести месяцев были осуществлены постоянные измерения течений на 10 глубинах от 25 до 1500 м на 17 заякоренных буйковых станциях, образовывавших крест размерами 200X200 км с центром в точке 16°ЗГ 14, 33°30 Ш, и был также исполнен ряд гидрологических съемок.
Так была внесена поправка в представление о не-возобновимости минеральных богатств. Пригодные ископаемые за исключением торфа и некоторых других обычных образований невозобновимы в отработанных месторождениях на досягаемой для человека глубине недр материков. Это и внятно - безвозвратно исчезли те физико-химические и иные данные в зоне месторождения, которые в дальнем прошлом геологической истории сотворили дорогие для человека минеральные образования. Другое дело добыча со дна присутствующего океана гранулированных руд. Мы можем их взять, а в сделавшей эти руды натуральной действующей лаборатории, какой является океан, не прекратятся процессы рудообразования.
Если гравитационные аномалии в свободном воздухе на материках и океанах не имеют принципиальных отличий, то в редукции Буге эта разница проявляется крайне приметно. Вступление поправки за могущество промежуточного слоя в океане приводит к приобретению высоких позитивных значений аномалий Буге, тем огромных, чем огромнее глубина океана. Данный факт обусловлен теоретическим нарушением при вступлении поправки Буге («засыпке» океана) природной изостазии океанической литосферы. Так, в гребневых зонах СОХ аномалия Буге составляет около 200мГал, для абиссальных океанических котловин - в среднем от 200 до 350 мГал. Бесспорно, что в аномалиях Буге отражаются генеральные черты рельефа дна океана в той степени, в какой они изостатически скомпенсированы, от того что стержневой взнос в аномалии Буге вносит именно теоретическая поправка.
Основными процессами, определяющими профиль окраины, возникшей у тылового края материка (пассивной окраины), являются примерно перманентные погружения, исключительно существенные в дистальной, приокеанической ее половине. Лишь отчасти они компенсируются накоплением осадков. Во времени окраина разрастается как в итоге вовлечения в прогибание все больше удаленных от океана континентальных блоков, так и в итоге образования сильной осадочной линзы на материковом подножии. Разрастание происходит основным образом за счет соседних участков океанского ложа и является следствием непрекращаю-щейся эрозии прилегающих к окраине районов материка, а также глубинных его областей. Это находит отражение не только в ненеиленизации суши, но и в смягчении, выравнивании рельефа в подводных участках зоны перехода. Происходит так сказать агградация: выравнивание поверхности переходных зон в областях с пассивным тектоническим режимом. Вообще говоря, эта склонность характерна для всякий окраины, впрочем в тектонически энергичных зонах она не реализуется в итоге орогенеза, складкообразования, роста вулканических построек.
В соответствии с особенностями морской воды температура ее даже на поверхности лишена крутых контрастов, присущных приземным слоям воздуха, и колеблется в пределах от -2°С (температура замерзания) до 29°С в открытом Океане (до 35,6°С в Персидском заливе). Но это объективно в отношении температуры воды на поверхности, обусловленной поступлением ясной радиации. В рифтовых зонах Океана на крупных глубинах открыты сильные гидротермы с температурой воды, находящейся под огромным давлением, до 250-300°С. И это не эпизодические излияния перегретых глубинных вод, а долго (даже по геологическим масштабам) либо же непрерывно существующие на дне Океана озерки сверхгорячей воды, о чем говорит их экологически уникальная бактериальная фауна, использующая для своего питания соединения серы. В этом случае амплитуда безусловного максимума и минимума температуры воды Океана составит 300°С, что в два раза превышает амплитуду предельно высоких и низких температур воздуха у земной поверхности.
Рассеивание вещества биострома распространяется на существенную часть толщи географической оболочки, а в атмосфере даже выходит за ее пределы. Жизнеспособные организмы найдены на высоте больше 80 км. В атмосфере нет самостоятельной жизни, но воздушная тропосфера - транспортер, переносчик на большое расстояние семян и спор растений, микробов, среда, в которой проводят существенную часть жизни многие насекомые и птицы. Рассеивание водно-поверхностного биострома распространяется на всю толщу океанических вод вплотную до донной пленки жизни. Дело в том, что глубже эвфотической зоны сообщества фактически лишены собственных продуцентов, энергетически они всецело зависимы от сообществ верхней зоны фотосинтеза и на этом основании не могут рассматриваться полновесными биоценозами в понимании Ю. Одума (М. Е. Виноградов, 1977). С нарастанием глубины биомасса и количество планктона стремительно уменьшаются. В батипелагиали в самых плодотворных районах океана биомасса не превышает 20-30 мг/м3 - это в сотни раз поменьше, чем в соответствующих районах на поверхности океана. Глубже 3000 м, в абиссопелагиали, биомасса и количество планктона экстраординарно низкие.








Добавить комментарий